Peptidoglycan
Peptidoglycan is the main constituent of the bacterial cell wall and is essential for cell viability. Accordingly, several classical antibiotics such as penicillin and vancomycin impair peptidoglycan biosynthesis to kill bacteria. Polymeric chains of GlcNAc and N-acetylmuramic acid (MurNAc) constitute the backbone and invariant core of peptidoglycan. Each disaccharide unit of GlcNAc(1-4) MurNAc comprise a peptidic side chain of five amino acids featuring rare D-amino acids. Whereas the carbohydrate moiety of peptidoglycan is invariant among bacteria, different peptide sequences occur across bacterial groups. For example in Escherichia coli the peptide is L-Ala~D-Glu~mA2pm~D-Ala~D-Ala, where mA2pm represents meso-diaminopimelic acid. In Gram-positive Staphylococcus aureus, the peptide includes a branch of five glycines linked to the middle lysine residue, yielding L-Ala~D-isoGlu~L-Lys[Gly]5~D-Ala~D-Ala.
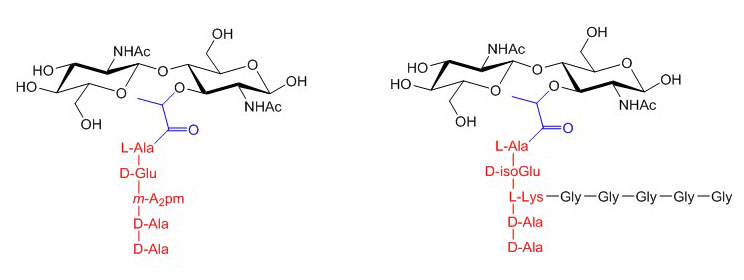
Figure X. Core structures of peptidoglycan from Escherichia coli (left) and Staphylococcus aureus (right). The lactyl group at C3 of MurNAc is in blue and the pentapeptide is in red. The Gly5 side-chain of Staphylococcus aureus is in black.
Although peptidoglycan is found in all bacteria containing a cell wall, its layout differs between Gram-negative and Gram-positive bacteria. Gram-negative bacteria have a double membrane and peptidoglycan is found as a thin layer on top of the inner cell membrane. Gram-positive bacteria lack an outer membrane and probably therefore require a much thicker layer of peptidoglycan to sustain the high internal osmotic pressure. Peptidoglycan is anchored to the inner membrane of Gram-negative bacteria through linkage to Braun’s lipoproteins and to the membrane of Gram-positive bacteria through cross-links to lipoteichoic acid.
Peptidoglycan biosynthesis begins in the cytoplasm by generating UDP-MurNAc through addition of a lactyl group to the C3 atom of UDP-GlcNAc. UDP-MurNAc is then linked to the pentapetide side chain and the resulting glycoconjugate is transferred to undecaprenol-P, yielding MurNAc-(pentapeptide)-PP-undecaprenol, which is also called lipid I. As next step, GlcNAc is transferred in a β1-4 linkage to MurNAc, thereby yielding lipid II. Units of lipid II are then translocated across the membrane through a yet unknown mechanism and polymerized by a transglycosylase enzyme in a reaction that releases undecaprenol-PP. The length of the GlcNAc-MurNAc polymer varies between bacterial families, usually within a range of 5 to 100 disaccharide units. After separation from the membrane bound lipid carrier, chains of peptidoglycan are cross-linked by bridging peptidic side-chains from different polysaccharide strands through a transpeptidase reaction. The efficiency of peptide cross-linking depends on the bacteria and growth conditions. Most bacteria reach 40 to 60% cross-linking, but Staphylococcus aureus achieves up to 90% cross-linking thank to the additional pentaglycine secondary chain.
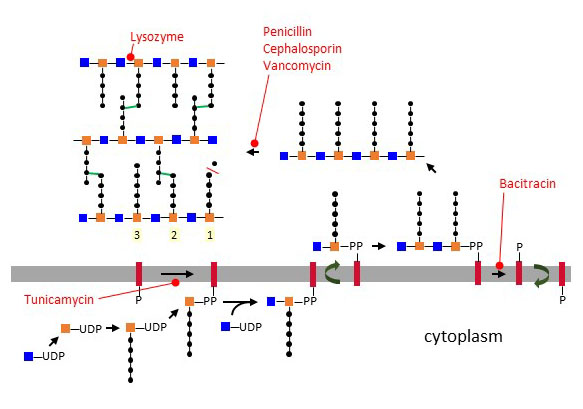
Figure X – Biosynthesis of peptidoglycan at the cell membrane and cross-linking in the periplasmic space. Reactions sensitive to antibiotics are marked in red.
Transglycosidases and transpeptidases belong to the class of penicillin-binding proteins (PBP). Bacterial genomes contain multiple PBP genes, which encode carboxypeptidases, transpeptidases, endopeptidases, and transglycosylases involved in peptidoglycan biosynthesis and remodeling at various locations of the cell wall. The cell wall is under constant remodeling to encompass changes of the cell architecture, such as during division. Peptidoglycan hydrolysis and biosynthesis are tightly coordinated to prevent any rupture of the cell wall. Several hypotheses have been proposed to explain the process. One hypothesis suggests that new layers are added closest to the cell membrane, whereas older layers are hydrolyzed at the top facing the extracellular space. Another concept, referred to as the “three-for-one” hypothesis, states that hydrolysis and biosynthesis takes place through a multi-enzyme complex that synthesizes three new strands for one old strand being hydrolyzed. β-Lactam antibiotics like penicillin block peptidoglycan biosynthesis by inhibiting transpeptidases. Vancomycin also interferes with peptidoglycan formation by binding D-Ala~D-Ala of the peptidic side-chains, thus preventing cross-linking. The antibiotic bacitracin works by inhibiting the dephosphorylation of undecaprenol-PP, which abolishes the recycling of the lipid carrier to the internal leaflet of the membrane for further biosynthesis of lipid I units. Interestingly, tunicamycin, which inhibits the first step of N-linked glycosylation in eukaryotes, also interferes with peptidoglycan formation by inhibiting the transfer of MurNAc to undecaprenol-P. In addition to the multitude of microbial antibiotics targeting peptidoglycans, animals also express the antibacterial protein lysozyme, which cleaves the glycosidic bond between GlcNAc and MurNAc.
Peptidoglycan remodeling liberates fragments into the environment, which are strong activators of the innate immune system. Peptidoglycan-recognition proteins are widely distributed among animals. For example, insects express a battery of at least 20 proteins involved in the cleavage, recognition, and presentation of peptidoglycan to immune cells. In mammals, the toll-like receptor-2 (TLR2)/CD14 complex recognizes polymeric peptidoglycan in the extracellular space. Peptidoglycan fragments are also recognized inside cells by the cytoplasmic NOD1 and NOD2 proteins. NOD1 recognizes L-Ala~D-Glu~mesoA2pm and NOD2 recognizes MurNAc~L-Ala~D-Glu. The mechanisms involved in the transfer of peptidoglycan fragments to the cytoplasm are however currently unknown. In addition to membrane-bound and cytoplasmic proteins, several secreted lectins, such as mannose-binding lectin and mouse RegIII, recognize GlcNAc-MurNAc polymers and contribute to bacterial elimination.