Milk oligosaccharides
The main constituent of milk carbohydrates, the disaccharide lactose, is only found in mammals. Lactose is produced in the lactating mammary gland by addition of b1-4 linked Gal to Glc. The b1-4 Gal-transferase enzyme B4GALT1, commonly found in the Golgi apparatus of all cell types, catalyzes this transfer reaction to Glc through binding to a-lactalbumin. Through interaction with a-lactalbumin, the acceptor specificity of B4GALT1 switches from GlcNAc to Glc, thereby enabling the biosynthesis of lactose. The pituitary hormone prolactin induces a-lactalbumin expression, which itself controls lactose production in the mammary gland epithelium.
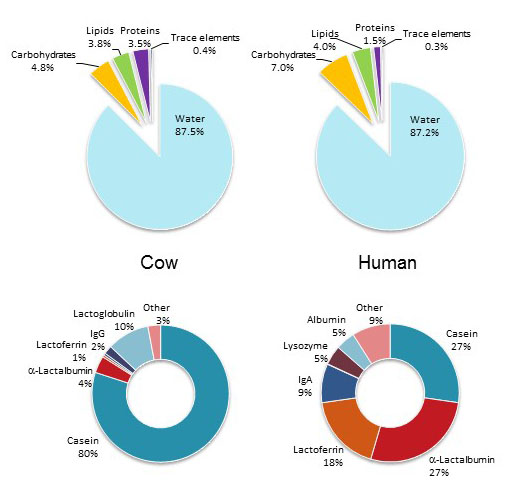
Lactose is not always a beneficial nutrient for human beings. Young children can digest lactose because they secrete lactase in their small intestine, but most adults are lactose-intolerant because lactase expression is turned off during childhood. About a third of human beings maintain lactase expression in adulthood thank to a mutation in the promoter region of the lactase gene, which appeared about 7500 years ago in Europe. Other mutations have been identified in the Middle East, West Africa, and South Asia conferring similar maintenance of lactase expression. The ability to use milk as nutrient provided a strong advantage to Neolithic farmers over hunter-gatherers and promoted their expansion across Europe, thereby further propagating the beneficial lactase gene polymorphism. Beyond lactose, milk contains a variable amount of lipids and proteins, of which some have antimicrobial activity such as immunoglobulins, lactoferrin, and lysozyme.
Figure X. Comparison of cow's milk and human milk composition based on main constiutent classes. The bottom diagrams show the relative amounts of individual proteins in cow's milk and human milk.
Golgi-localized glycosyltransferases elongate lactose further, yielding a complex mixture of oligosaccharides ranging from trisaccharides to structures consisting of 20 or more monosaccharides. To date, structural information on milk oligosaccharides have been collected in nearly 90 mammalian species. This survey revealed a great structural diversity with some types of milk including only few oligosaccharides as found in rodents, and others like human milk including up to 200 distinct oligosaccharides. All primates also show a large diversity in milk oligosaccharides, although reaching only up to half the diversity found in human beings. Oligosaccharide concentrations are highest (10-20 g/l) in the first milk called colostrum, which is delivered in the first days after birth. Over the following weeks, oligosaccharide concentrations decrease and stabilize around 5-10 g/l. Milk oligosaccharides are built up from the five monosaccharides Glc, Gal, GlcNAc, Fuc, and Sia. The most abundant oligosaccharides in human milk are the trisaccharide 2’-O-fucosyllactose (2-3 g/l) and the pentasaccharide lacto-N-fucopentaose I (1.0-1.8 g/l). By comparison, lactose concentration reaches about 70 g/l in human milk.
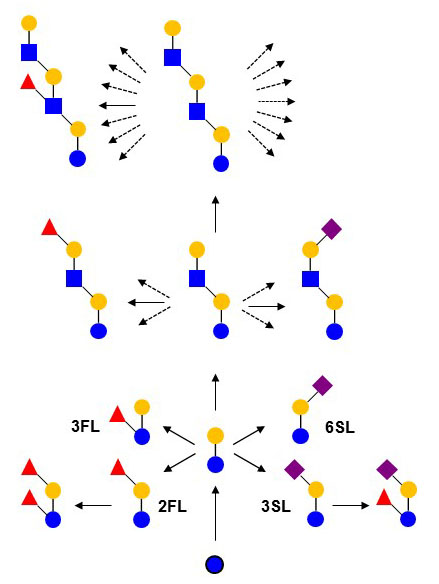
Figure X. Biosynthesis of milk oligosaccharides through the extention of lactose by Golgi-resident glycosyltransferases. 2FL: 2’-O-fucosyllactose, 3FL: 3’-O-fucosyllactose, 3SL: 3’-O-sialyllactose, 6SL: 3’-O-sialyllactose.
The composition of milk oligosaccharides is under genetic control and shows a strong individual variability between lactating mothers. Besides such inter-individual variations, the concentration of specific oligosaccharides vary over the course of lactation with some oligosaccharides decreasing and others increasing over time. Common carbohydrate antigens like blood group A and B are not found on human milk oligosaccharides, but they do occur in the milk of other mammals. By contrast, human milk oligosaccharides expose multiple Lewis antigens, such as LeA, LeB, LeX, LeY, Sialyl LeA, and Sialyl LeX. The Secretor (Se) status affects the fucosylation of human milk oliogsaccharides. Accordingly, Se- mothers have no a1-2 linked Fuc on their milk oligosaccharides.
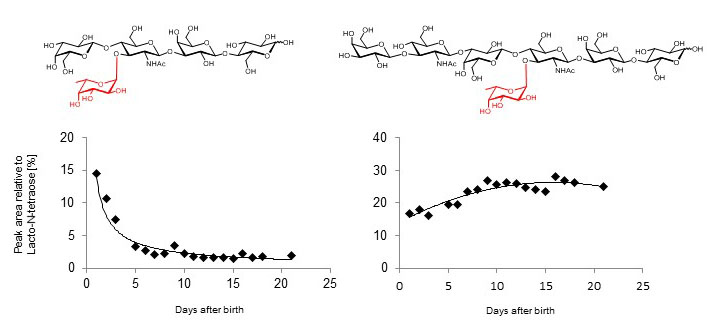
Figure X. Concentration of lacto-N-fucopentaose III (left panel) and fucosyl-lacto-N-hexaose (right panel) in the breast milk of a single mother during the first 20 days of lactation.
{kind=link}
{kind=link}
Milk oligosaccharides are neither cleaved, nor absorbed in the intestinal tract, thus they are not used as a nutrients by the suckling infant. The question of their functional relevance remains largely speculative, although a connection between breastfeeding and intestinal bacteria has been recognized already more than a century ago. Around 1900, Escherich, Moro, Tissier described differences in fecal bacteria between breastfed and bottle-fed infants. By 1930, Schönfeld demonstrated that the previously described bifidus factor affecting the growth of intestinal bifidobacteria was in fact a carbohydrate fraction of breast milk. This fraction was first called gynolactose and later found to consist of various fucosylated structures. Several families of intestinal bacteria, such as Bacteroidaceae and Bifidobacteriaceae, express glycosidases that break down milk oligosaccharides and then utilize monosaccharides as source of energy. In this respect, milk oligosaccharides contribute to shaping the intestinal microbiota.
Through their similarity to cellular glycans, milk oligosaccharides can also function as soluble receptors for intestinal bacteria and viruses. For example, sialylated milk oligosaccharides can neutralize influenza virus through binding to hemagglutinin. Similarly, fucosylated milk oligosaccharides also bind to several strains of norovirus, thereby preventing the docking of the virus to cellular glycan receptors. Milk oligosaccharides also inhibit the adhesion of bacteria such as Campylobacter jejuni, Listeria monocytogenes, and Streptococcus pneumonia to the intestinal epithelium. Furthermore, milk oligosaccharides protect against bacterial enterotoxins such as cholera toxin by competing with cell-bound glycan receptors.
Milk oligosaccharides can also bind to endogenous lectins, such as siglecs and galectins, which are expressed on mucosal immune cells. It is therefore likely that oligosaccharides participate to the maturation of intestinal immunity and recognition of carbohydrate antigens taken up orally. The beneficial effect of breast milk on infantile mortality was noticed by the beginning of the 20th century. Nowadays, bottle-fed infants still have a five to ten-fold increased risk compared to breast-fed infants of developing necrotizing enterocolitis, which affects about 5% of premature infants.