Viral glycosylation
Glycosylation occurs not only in all domains of life, but also in viruses. The role of host-derived glycans in virus infection, replication, and immune evasion will be discussed later in the chapter dedicated to infections and toxins. Viruses have learnt to exploit host glycosylation in different ways to ensure their propagation and survival as much inside as outside of their host. Viruses such as cytomegalovirus and HTLV1 induce the expression of the host fucosyltransferase FUT7 gene, which increases the presentation of sialyl-LewisX at the surface of infected cells and thereby promotes the infiltration of infected leukocytes through tissues.
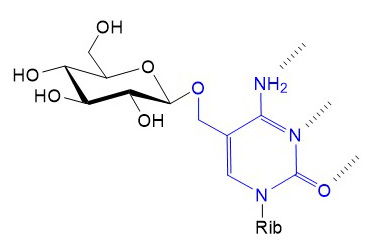
Several viral genome include glycosyltransferase genes, which are expressed during virus replication. Most of these viral glycosyltransferases modify cellular targets to gain advantages during replication or post-infective propagation of virus particles. Glycosylation is not just a strategy applied by large viruses with complex genomes. In fact, bacteriophages T2, T4 and T6 express glycosyltransferases linking Glc to hydroxy-methyl-cytosine, which when incorporated into the phage genome prevents its digestion by bacterial restriction enzymes.
Figure X. Glycosylation of hydroxy-methyl-cytosine by bacteriophage glucosyltransferases. Hydrogen bonds between cytosine and guanine are marked with dashed lines. Rib: (deoxy)ribose.
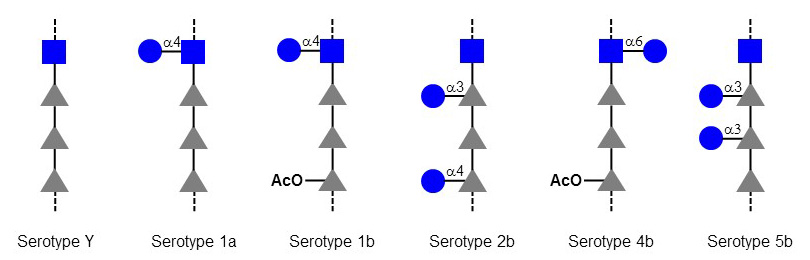
The so-called seroconverting phages of Shigella flexneri use a different approach. These lysogenic phages integrate their DNA into the bacterial chromosome and express Glc-transferases and O-acetyltransferases, which modify O-antigen polysaccharides thereby changing the serotype of infected Shigella. The alteration of O-antigen prevents the infection of bacteria by other phages using O-antigen determinants as receptor. Seroconversion also contributes to the evasion of Shigella from neutralizing antibodies recognizing the original O-antigen.
Figure X. O-Antigen modifications of Shigella flexneri by seroconverting bacteriophages. The Glc and O-acetyl (AcO) residues are transferred by bacteriophage genes.
Some baculoviruses exploit an interesting target to maximize their propagation. Baculoviruses infecting Lepidoptera larvae express a Glc-transferase adding Glc to the molting hormone ecdysone, which inactivates the hormone and thereby prevents the metamorphosis of the caterpillar host. The inactivation of ecdysone also influence the behavior of the infected caterpillar, which remains on leaves instead of burrowing into the soil in preparation for pupation. The death and lysis of infected caterpillars on leaves releases baculovirus particles exactly where new hosts will feed and become infected.
The rationale for the expression of glycosyltransferases by viruses is not always as clear as in the cases of baculoviruses and bacteriophages. For example, bovine herpes virus BHV4 expresses a core-2 β1-6 GlcNAc-transferase, which activity is not essential for virus replication. The genome of the South American myxoma virus includes a α2-3 Sia-transferase, which glycosylates a SERP1 protein also encoded by the virus genome. The role of this viral sialylated SERP1 serpin is unclear, but may be related immune suppression. Serpins are protease inhibitors and the most prominent members of this protein family, α1-antitrypsin and antithrombin, are involved in the regulation of coagulation and inflammatory cascades. Besides myxoma virus, other leporypoxviruses such as Shope fibroma virus also express a sialyltransferase, suggesting a general advantage provided by this enzyme to the viruses despite its dispensability for virus replication.
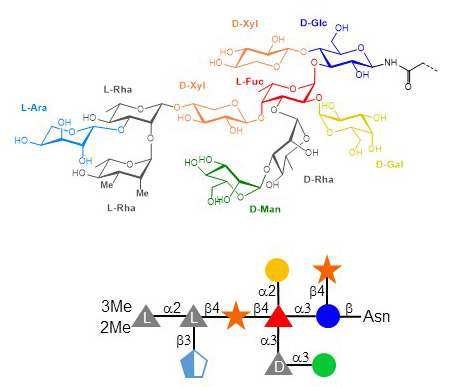
The examples outlined so far consisted of single glycosyltransferase genes in viral genomes. Some viruses however express multiple glycosyltransferase genes and even genes required for the biosynthesis of nucleotide-activated sugar substrates. Such viruses that are often coined as giant viruses belong to the group of nucleocytoplasmic large DNA viruses, which comprises Poxviridae, Asfarviridae, Iridoviridae, Ascoviridae, Phycodnaviridae, Megaviridae, and Marseilleviridae. The first large DNA virus described to include a glycosylation system was Paramecium bursaria chlorella virus PBCV-1. This icosahedral virus of 200 nm diameter infects chlorella algae, which are photosynthetic endosymbionts of the protozoan Paramecium bursaria. The PBCV-1 DNA genome of 330 kbp includes several genes usually not found in viruses, such genes encoding tRNA, DNA repair enzymes, DNA methyltransferases, and seven glycosyltransferases. In fact, these glycosyltransferases are still putative since their enzymatic activities have not been demonstrated yet. The PBCV-1 genome also encodes enzymes mediating the formation of GDP-Fuc and GDP-Rha. The major capsid protein of PBCV-1 carries N- and O-glycans produced by virally-encoded glycosyltransferases. The main N-glycans represent complex structures of nine to ten monosaccharides featuring a rare triple-substituted Fuc residue. The attachment site of the four N-glycans to the Vp54 capsid protein is also unique, as the context of the Asn acceptor does not match the classical consensus sequence Asn-X-Ser/Thr of N-glycosylation. The significance of N-glycans on Vp54 is unclear; all monosaccharides beyond the core Xyl(β1-4)Glc are non-essential since mutant viruses displaying truncated N-glycans are stable and infectious. PBCV-1 and related chlorella viruses also express hyaluronan and chitin synthesizing genes. Here too, the functional relevance of these polysaccharides for the viruses is unknown.
Figure X. N-glycan of PBCV-1 Vp54 capsid protein. Some N-glycoforms lack the β3-linked Ara residue.
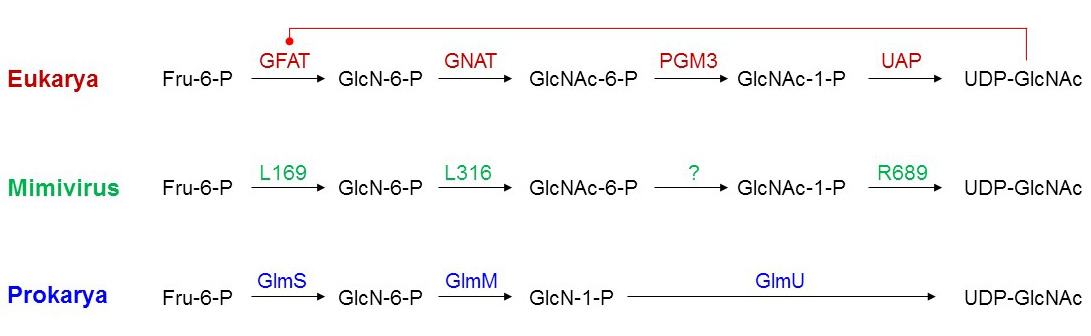
Even more complex glycosylation systems than in PBCV-1 exist in Megaviridae, which consist of mimivirus, moumouvirus, megavirus and the like. These giant viruses have genomes exceeding the Mbp mark and encompassing more than 1000 genes. Each type of Megaviridiae express on average between 10 and 15 putative glycosyltransferases. Structurally, these viral glycosyltransferase candidates are similar to archaeal, bacterial and eukaryotic glycosyltransferases. To date, a single mimivirus glycosyltransferase has been shown to glycosylate collagen-like proteins, which are also encoded by the mimivirus genome. Genes encoding complete biosynthetic pathways for nucleotide-activated sugars have been characterized, thereby outlining the production of exotic sugars that are usually only found in few prokaryotes. These nucleotide-activated sugars include UDP-viosamine (4-amino-4,6-dideoxy-D-Glc), UDP-RhaNAc, but also the ubiquitous UDP-GlcNAc. The examination of the UDP-GlcNAc biosynthesis pathway in mimivirus provided interesting clues about the evolution of glycosylation systems in giant viruses. The biosynthesis of UDP-GlcNAc in mimivirus proceeds through the same steps as the eukaryotic pathway but some individual mimivirus enzymes share properties with their bacterial counterparts. For example, eukaryotic glutamine-fructose-6-phosphate transaminase (GFAT) is inhibited by UDP-GlcNAc but the mimivirus enzyme, like the bacterial glucosamine-6-P synthase GlmS, is unaffected by UDP-GlcNAc levels. The comparison between giant virus, prokaryotic and eukaryotic pathways indicate that viruses have not just captured bacterial or eukaryotic genes into their genomes, but developed intermediate strategies suggesting a parallel evolution to the kingdoms of life. Glycosylation pathways are found in all giant viruses identified to date, suggesting an important contribution of glycoconjugates in virus replication and propagation.
Figure X. UDP-GlcNAc biosynthesis pathway in eukaryotes, prokaryotes and mimivirus. Eukaryotic GFAT is inhibited by UDP-GlcNAc.
{kind=link}
{kind=link}
{kind=link}
{kind=link}